
Scott Lab
Primate trophic adaptation, trait evolution, and diversification
The Scott Lab investigates questions related to the evolution of the primate chewing apparatus and the factors that have shaped primate biodiversity. Current lab projects are aimed at understanding the interplay between ecology, anatomy, and diversification in living primates. The results of these studies provide a framework for reconstructing primate evolutionary history and inferring feeding adaptations in extinct primates, including fossil members of our own lineage.
Dietary influences on primate tooth size
Humans are unusual in comparison to other primates in having small teeth and jaws relative to body size. The human fossil record indicates that different teeth experienced evolutionary size reduction at different times over the past 7 million years. Molar and premolar reduction has been linked to our use of cultural practices to prepare food before the first bite, reducing the amount of chewing required. However, some of our ancient relatives evolved very large molars and premolars before going extinct. They also had massive jaws combined with skull shapes indicating the presence of correspondingly massive jaw muscles. What were they eating? Since we cannot directly observe their behavior, we must make inferences using links between form and function found in living primates.

The upper teeth and jaws of two extant primates, illustrating differences in relative molar and premolar size. Both species are lemurs from Madagascar. Eulemur fulvus (common brown lemur) eats a diet of fruit and leaves, whereas Hapalemur griseus (lesser bamboo lemur) specializes on bamboo parts, which are mechanically challenging to process. The two specimens are scaled to the same jaw length to illustrate the relatively larger molars and premolars of Hapalemur (yellow lines).
Primate macroevolution and trait evolution
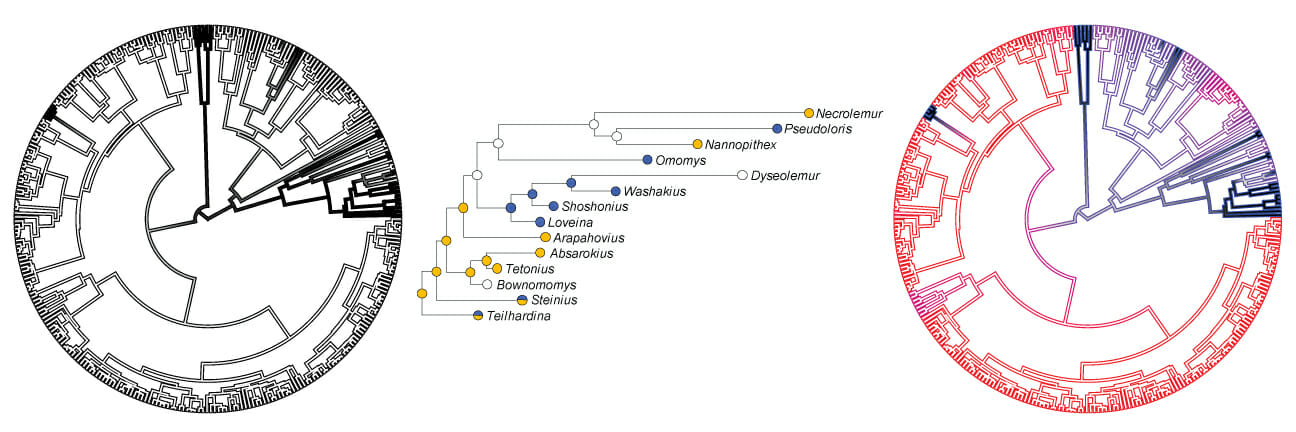
Some primate lineages contain many species, while others contain only a few. Some traits are evenly distributed across primates, while others are clumped in particular groups. What factors have played a role in structuring primate biodiversity? Why are certain traits common in some groups but rare in others? What do the patterns observed in living species tell us about primate biology? These questions fall within the realm of macroevolutionary analysis: the study of rates of speciation, extinction, and trait evolution and how variation in those rates has shaped the tree of life and the distribution of traits at its tips.

Private phylogenetic tree showing the distribution of herbivory and insectivory among small-bodied species at the tips of the tree. Insectivory is found only in small-bodied species, probably because large-bodied primates (unmarked tips) have a difficult time acquiring enough insects to meet their daily caloric requirements. Note how insectivory is unevenly distributed among small-bodied primates, being far more common in lineages that are nocturnal. What is the reason for this pattern?
Nalley TK, Scott JE, Ward CV, Alemseged Z, 2019. Comparative morphology and ontogeny of the thoracolumbar transition in great apes, humans, and fossil hominins. Journal of Human Evolution 134, 102632.
Scott JE, 2019. Macroevolutionary effects on primate trophic evolution and their implications for reconstructing primate origins. Journal of Human Evolution 133, 1–12.
Scott JE, 2018. Reevaluating cases of trait-dependent diversification in primates. American Journal of Physical Anthropology 167, 244–256.
Scott JE, Campbell RM, Baj LM, Burns MC, Price MS, Sykes JD, Vinyard CJ, 2018. Dietary signals in the premolar dentition of primates. Journal of Human Evolution 121, 221–234.
Franks EM, Scott JE, McAbee KR, Scollan JP, Eastman MM, Ravosa MJ, 2017. Intracranial and hierarchical perspective on dietary plasticity in mammals. Zoology 124, 30–41.
Lewton KL, Scott JE, 2017. Ischial form as an indicator of bipedal kinematics in early hominins: a test using extant anthropoids. The Anatomical Record 300, 845–858.
Franks EM, Holton NE, Scott JE, McAbee KR, Rink JT, Pax KC, Pasquinelly AC, Scollan JP, Eastman MM, Ravosa MJ, 2016. Betwixt and between: intracranial perspective on zygomatic arch plasticity and function in mammals. The Anatomical Record 299, 1646–1660.
Ravosa MJ, Menegaz RA, Scott JE, Daegling DJ, McAbee KR, 2016. Limitations of a morphological criterion of adaptive inference in the fossil record. Biological Reviews 91, 883–898.
Ravosa MJ, Scott JE, McAbee KR, Veit AJ, Fling AL, 2015. Chewed out: an experimental link between food mechanical properties and repetitive loading of the masticatory apparatus in mammals. PeerJ 3, e1345.
Scott JE. Lost and found: the third molars of Callimico goeldii and the evolution of the callitrichine postcanine dentition, 2015. Journal of Human Evolution 83, 65–73.
Scott JE, McAbee KR, Eastman MM, Ravosa MJ, 2014. Teaching an old jaw new tricks: diet-induced plasticity in a model organism from weaning to adulthood. Journal of Experimental Biology 217, 4099–4107.
Scott JE, 2014. Cranial size variation and lineage diversity in early Pleistocene Homo. Evolution 68, 909–915.
Scott JE, McAbee KR, Eastman MM, Ravosa MJ, 2014. Experimental perspective on fallback foods and dietary adaptations in early hominins. Biology Letters 10, 20130789.
Doney E, Krumdick LA, Diener JM, Wathen CA, Chapman SE, Stamile B, Scott JE, Ravosa MJ, Van Avermaete T, Leevy WM, 2013. 3D printing of preclinical x-ray computed tomographic data sets. Journal of Visualized Experiments 73, e50250.
Scott JE, 2012. Molar size and diet in the Strepsirrhini: implications for size-adjustment in studies of primate dental adaptation. Journal of Human Evolution 63, 796–804.
Scott JE, Lack, JB, Ravosa MJ, 2012. On the reversibility of mandibular symphyseal fusion. Evolution 66, 2940–2952.
Scott JE, Hogue AS, Ravosa MJ, 2012. The adaptive significance of mandibular symphyseal fusion in mammals. Journal of Evolutionary Biology 25, 661–673.
Scott JE, 2011. Folivory, frugivory, and postcanine size in the Cercopithecoidea revisited. American Journal of Physical Anthropology 146, 20–27.
Royer DF, Lockwood CA, Scott JE, Grine FE, 2009. Size variation in the early human mandibles and molars from Klasies River, South Africa: Comparison with other middle and late Pleistocene assemblages and with modern humans. American Journal of Physical Anthropology 140, 312–323.
Scott JE, Schrein CM, Kelley J, 2009. Beyond Gorilla and Pongo: Alternative models for evaluating variation and sexual dimorphism in fossil hominoid samples. American Journal of Physical Anthropology 140, 253–264.
Scott JE, Marean CW, 2009. The Paleolithic hominin remains from Eshkaft-e Gavi (southern Zagros Mountains, Iran): description, affinities, and evidence for butchery. Journal of Human Evolution 57, 248–259.
Orr CM, Delezene LK, Scott JE, Tocheri MW, Schwartz GT, 2007. The comparative method and the inference of venom-delivery systems in fossil mammals. Journal of Vertebrate Paleontology 27, 541–546.
Scott JE, Stroik LK, 2006. Bootstrap tests of significance and the case of humanlike skeletal-size dimorphism in Australopithecus afarensis. Journal of Human Evolution 51, 422–428.
Scott JE, Lockwood CA, 2004. Patterns of tooth crown size and shape variation in great apes and humans and species recognition in the hominid fossil record. American Journal of Physical Anthropology 125, 303–319.
JEREMIAH SCOTT, PHD

College of Osteopathic Medicine of the Pacific
Assistant Professor of Anatomy